Das süddeutsche Molassebecken
 This text, unless otherwise expressly stated, is licensed under a Creative Commons Attribution-Share Alike 3.0 Germany License.
This text, unless otherwise expressly stated, is licensed under a Creative Commons Attribution-Share Alike 3.0 Germany License.Inhaltsverzeichnis
- Untergrund des westlichen Molassebeckens
- Ammoniten-Zonen des westlichen Molassebeckens
- Die Ostracodenfaunen des westlichen Molassebeckens
- Beschreibung einiger Ostracoden aus dem Molassebecken
- 4.1 Ostracode 1
- 4.2 Ostracode 2
- 4.3 Ostracode 3
- 4.4 Ostracode 4
- Literatur
Das süddeutsche Molassebecken
1. Untergrund des westlichen Molassebeckens
Die Bohrungen Fronhofen 8, Harresheim 1 und Markdorf 3 sind für dieses Gebiet repräsentativ. Im Untergrund des westlichen Molassebeckens ist der gesamte Unterjura nur lückenhaft vorhanden und meistens kondensiert. Vermutlich führte die Vindelizische Schwelle zur Ablagerung von geringmächtigen Unterjurafolgen. Die Schüttungen von Sanden sowie die Bildung von Kalkooiden deuten auf einen Ablagerungsraum in Küstennähe hin. Die Lage über der oberen Wellenbasis führt zur Erosion und Resedimentation der älteren Schichtenfolgen. Die auftretenden Sande stammen vermutlich von der Vindelizischen Schwelle bzw. von der Alemannischen Insel. Da in Bohrungen Makrofossilien, vor allem Ammoniten, ausgesprochen selten sind, ist die stratigraphische Zuordnung der Schichtenfolgen nicht in jedem Fall durch entsprechende Leitfossilien gesichert. Die in den Bohrprotokollen angegebene stratigraphische Zuordnung erfolgte daher im wesentlichen nach lithologischen Gesichtspunkten. Sie wurde entsprechend der Probennahme und Profilbeschreibung zu Grunde gelegt.
Die Arietenkalke im Liegenden bestehen aus harten, dunklen, zum Teil sandigen Mergelkalksteinen, stellenweise auch aus Grobsandsteinen (Bohrung Harresheim 1). In den Profilen Harresheim 1 und Markdorf 3 treten in den oberen Mergelkalken Kalkooide auf. Die nachfolgenden Obtusustone bestehen aus dunkelgrauen, siltigen Tonen, in die je nach Entfernung zur Küste Sandstein‑ und Mergelkalksteinlagen eingeschaltet sind. In dem Profil Markdorf 3 treten in den tieferen Obtusustonen noch Kalkoolithe auf. Eine beta-Kalkbank ist nicht erkennbar, ihr Vorhandensein wurde jedoch von SCHNEIDER (1962) nicht ganz ausgeschlossen. Der Übergang zu den Numismalismergeln tritt lithologisch nicht in Erscheinung, er wird nur gelegentlich durch schwarze, kristalline Gryphaeenbänke markiert. Auch in den tieferen Numismalismergeln können Kalkooide auftreten. Numismalismergel und Obtusustone zusammen erreichen im westlichen Molassebecken eine Mächtigkeit von nur 6–7 ⅿ.
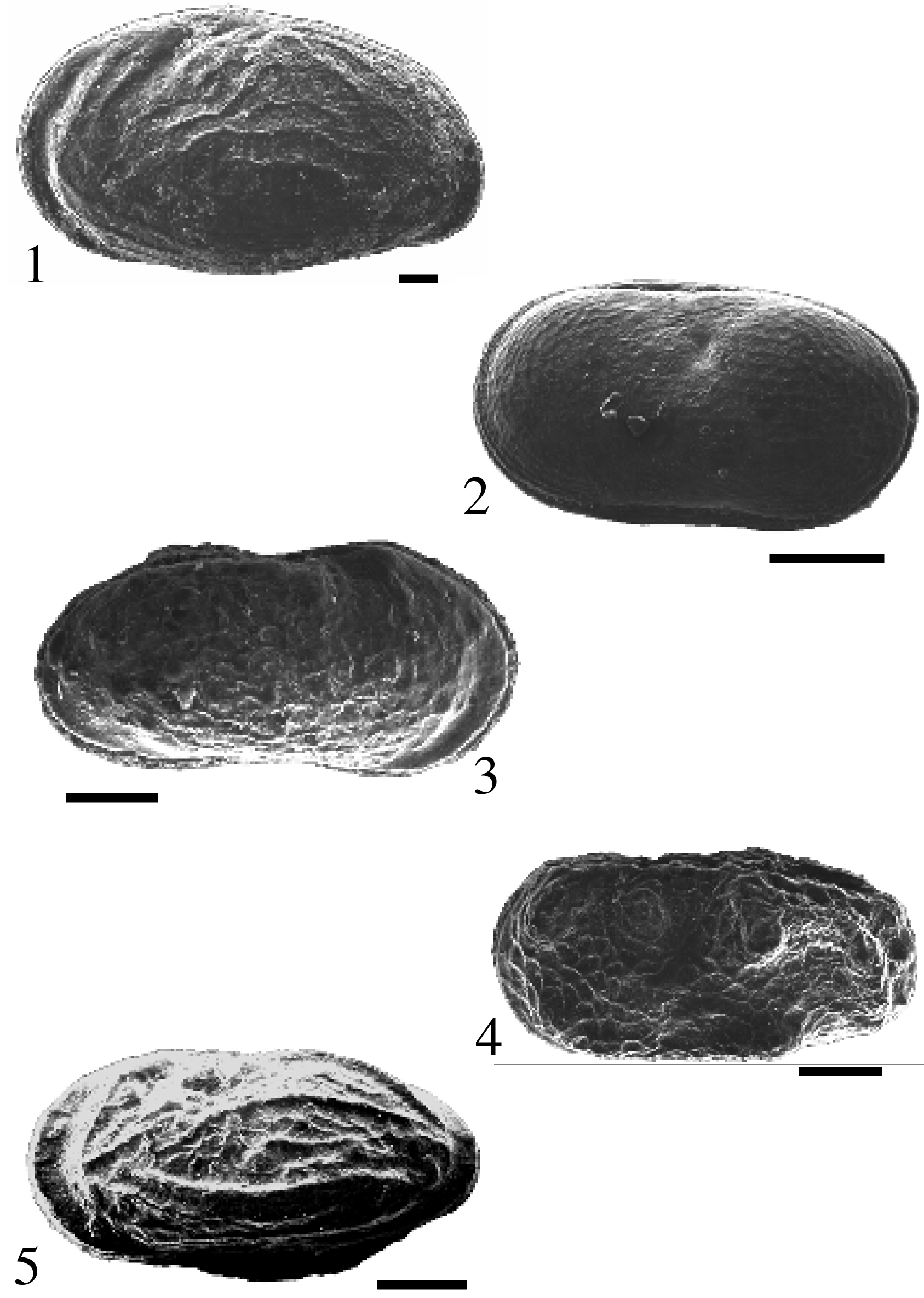
Schautafel – Figuren 1–5:
Der Maßstab entspricht jeweils 0,1 mm:
Fig. 1 : Ostracode 1, linke Klappe, Lateralansicht, Externseite; Bohrung Markdorf 3, Probe Nr. 9, vermutlich Sinemurium; BLNr. BEHER MA/9⁄184.
Fig. 2: Cytherella concentrica FIELD, Karapax, Lateralansicht, linke Externseite; Bohrung Markdorf 3, Probe Nr. 9, vermutlich Sinemurium; BLNr. BEHER MA/9⁄182.
Fig. 3: Ostracode 2, Karapax, Lateralansicht, rechte Externseite; Bohrung Markdorf 3, Probe Nr. 9, vermutlich Sinemurium; BLNr. BEHER MA/9⁄181
Fig. 4: Ostracode 3. Karapax, Lateralansicht, linke Externseite; Bohrung Harresheim 1, Probe Nr. 4, vermutlich Sinemurium oder Pliensbachium; BLNr. BEHER HA/4⁄177.
Fig. 5: Ostracode 4, Karapax, Lateralansicht, linke Externseite; Bohrung Markdorf 3, Probe Nr. 9, vermutlich Sinemurium; BLNr. BEHER MA/9⁄183.
Alle Ostracoden befinden sich in der Belegsammung zur Dissertation Beher (2001) und werden im Staatlichen Museum für Naturkunde Stuttgart aufbewahrt.
2. Ammoniten-Zonen des westlichen Molassebeckens
Im westlichen Molassebecken ist eine ammonitenstratigraphische Gliederung nicht möglich, da keine Ammonitenfunde vorliegen (vgl. SCHNEIDER, 1962).
3. Die Ostracodenfaunen des westlichen Molassebeckens
3.1. Die Ostracodenfaunen in Profil Harresheim 1
")
Bei dem Profil Harresheim handelt es sich um die Bohrung Harresheim 1, welche in den sechziger Jahren zur Erdölerkundung niedergebracht wurde. Aus diesem Grund ist die Bohrung nicht vollständig gekernt, und es konnte nur ein Teil des Materials bearbeitet werden. Die stratigraphische Gliederung der Gesteinsabfolge erfolgte anhand eines Gamma-Logs und es ergeben sich Unterschiede zwischen der lithostratigraphischen und biostratigraphischen Gliederung. Nach den Angaben des Gamma-Logs wurde das Profil ab 1 ⅿ unterhalb des Überganges der Arietenkalke in die Obtusutone/Numismalismergel (Anm.: eine Trennung dieser Folgen ist mit dem Gamma-Log nicht möglich.) bis 5 ⅿ unterhalb der Basis der Amaltheentone beprobt. Die oberen Meter lagen nur als Spülproben vor. Die Proben aus dem in der Bohrung Harresheim 1 als Sandstein ausgebildeten Arietenkalk erwiesen sich als fossilleer. Mit Beginn der Obtusustone/Numismalismergel setzen erste Ostracodenarten ein. Neben bekannten Arten wie Anchistrocheles? tuningensis, Fabalicypris praelonga, Donzocythere convergens, den Durchläuferarten Isobythocypris tatei und Ledahia sp., treten unbekannte Formen, hier mit Ostracode 1 und Ostracode 3 bezeichnet, auf. Kurz darauf folgen weitere nur zum Teil bekannte Ostracodenarten. Neben Cytherella concentrica, Paracypris sp. , Paradoxostoma? sp., Bairdiacypris sp., erscheinen auch Ledahia sp., Paracypris? redcarensis, Ostracode 2 und Ostracode 4. Im höheren Teil der Profilstrecke geht die Zahl der Ostracoden stark zurück, das Top fossilfrei.
3.1.1. Biostratigraphische Gliederung der Bohrung Harresheim 1
Für eine mögliche biostratigraphische Giederung des Profils können nur sehr wenige der auftretenden Arten herangezogen werden:
1. Donzocythere convergens (GRÜNDEL, 1964): Im Zürcher Weinland und auf der mittleren Schwäbischen Alb konnte diese Art im höheren Unter-Sinemurium und tiefem Ober-Sinemurium nachgewiesen werden, in der Literatur wird sie zumeist aus dem Unter-Sinemurium (Frankreich: DONZE, 1968) und denm obersten Hettangium bis unterstes Sinemurium hineinreichend (Celtic Sea: AINSWORTH, 1989) beschrieben.
2. Anchistrocheles? tuningensis BEHER, 2004: Diese Art wurde bisher nur aus Süddeutschland und Nordschweiz beschrieben. Dort ist ihr Vorkommen auf das Unter-Sinemurium und tieferes Ober-Sinemurium beschränkt (BEHER, 2004)
3. Cytherella concentrica FIELD, 1966: Aus Dorset/Engand wird diese Art aus dem Hettangium und Unter-Sinemurium beschrieben, HARLOFF (1993) bezeichnet eine aus dem Unter-Pliensbachium der Schwäbischen Alb stammende ähnliche Art mit Cytherella sp. A, und nach BRAND (1990) kommt die Art noch im Ober-Bathonium von Nordwestdeutschland vor. In der Langenbrückener Senke, im Randen/Zürcher Weinland und im Wutachgebiet setzt diese Art jedoch erst im höheren Ober-Sinemurium ein, in Profil Aselfingen reicht sie bis ins Unter-Pliensbachium hinein (BEHER, 2004). Im Bereich der mittleren Schwäbischen Alb und in der Nordschweiz ist sie auf das tiefere Ober-Sinemurium beschränkt.
4. Fabalicypris praelonga DONZE, 1966: In Frankreich ist diese Art von Hettangium bis ins tiefe Sinemurium hinein verbreitet (DONZE, 1966, 1985). In Süddeutschland und Nordschweiz ist sie überwiegend auf das tiefere Ober-Sinemurium beschränkt, einzig im Bereich der östlichen Schwäbischen Alb gibt es Funde aus der Grenze Ober-Sinemurium/Unter-Pliensbachium (BEHER, 2004).
5. Die hier mit Ostracode 1 bis Ostracode 4 bezeichneten Arten sind bisher unbekannt und bei der biostratigraphischen Gliederung des Bohrprofils nicht hilfreich. Das Auftreten von Anchistrocheles? tuningensis und Donzocythere convergens könnte den Beginn des Ober-Sinemurium kennzeichnen. Da die übrigen Arten auch im Bereich der Grenze Ober-Sinemurium/Unter-Pliensbachium vorkommen, können sie zur Gliederung nicht herangezogen werden.
3.2 Die Ostracodenfaunen in Profil Markdorf 3
") Bei dem Profil Markdorf handelt es sich um die Bohrung Markdorf 3 und hier stellt sich die Sachlage ähnlich wie bei Profil Harresheim dar. So wurde die Gesteinsabfolge ebenfalls nach dem Gamma-Log gegliedert und die Einheiten Obtusustone und Numismalismergel zusammengefaßt. Es wurde der oberste Abschnitt des Arietenkalks und ein mit Lias beta-gamma bezeichneter Abschnitt beprobt. Die nächsthöhere, mit Lias gamma-zeta bezeichnete Einheit lag nur als Spülprobe vor und wurde nicht bearbeitet.
Bei dem Profil Markdorf handelt es sich um die Bohrung Markdorf 3 und hier stellt sich die Sachlage ähnlich wie bei Profil Harresheim dar. So wurde die Gesteinsabfolge ebenfalls nach dem Gamma-Log gegliedert und die Einheiten Obtusustone und Numismalismergel zusammengefaßt. Es wurde der oberste Abschnitt des Arietenkalks und ein mit Lias beta-gamma bezeichneter Abschnitt beprobt. Die nächsthöhere, mit Lias gamma-zeta bezeichnete Einheit lag nur als Spülprobe vor und wurde nicht bearbeitet.
An der Basis des untersuchten Abschnittes treten neben Cytherella concentrica auch Bairdiacypris sp., Ledahia sp. und Ostracode 1 auf. Der Übergangsbereich von Arietenkalk in die Obtusustone/Numismalismergel hinein ist fossilleer. Erst 2 Meter über der Grenze setzen Cytherella concentrica und Ledahia sp. wieder ein. Desweiteren treten Paracypris sp. und Ostracode 1 dicht gefolgt von Fabalicypris praelonga, Isobythocypris tatei, Polycope sp., Ostracode 2, Ostracode 3 und Ostracode 4 auf. Etwas später erscheinen Bairdia molesta, Donzocythere convergens, Ogmoconcha sp. und Polycope cincinnata. Im letzten Abschnitt des Profils etwa 2 ⅿ unter der Grenze Obtusustone-Numismalismergel/Amaltheentone kommen Bairdia fortis, Cardobairdia liassica, Ogmoconchella sp. 1 und Pontocyprella fabaeformis dicht gefolgt von Bairdiacypris sp. vor. Den Abschluß bilden schließlich Bairdiacypris anisica brevis, Cytherelloidea lacertosa, Ogmoconcha aff. amalthei und Polycope cerasia.
3.2.1. Biostratigraphische Gliederung der Bohrung Markdorf 3
Für eine mögliche biostratigraphische Gliederung des Profils können nur sehr wenige der auftretenden Arten herangezogen werden:
1. Donzocythere convergens (GRÜNDEL, 1964): Im Zürcher Weinland und auf der mittleren Schwäbischen Alb konnte diese Art im höheren Unter-Sinemurium und tiefem Ober-Sinemurium nachgewiesen werden, in der Literatur wird sie zumeist aus dem Unter-Sinemurium (Frankreich:DONZE, 1968) und denm obersten Hettangium bis unterstes Sinemurium hineinreichend (Celtic Sea: AINSWORTH, 1989) beschrieben.
2. Cytherelloidea lacertosa (APOSTOLESCU 1959): In Süddeutschland kommt diese Art bisher im höheren Ober-Sinemurium und im Unter-Pliensbachium vor (CONTINI & PARIWATVORN, 1964; WALTSCHEW, 2000; BEHER; 2004). In Frankreich wird ihre Reichweite vom Ober-Sinemurium bis tiefstes Unter-Piensbachium (OERTLI, 1963; DONZE, 1985) angegeben bzw. im Pariser Becken erscheint sie bereits im tieferen Ober-Sinemurium und reicht in das höhere Unter-Pliensbchium hinein (APOSTOLESCU, 1959).
3. Cytherella concentrica FIELD, 1966: Aus Dorset/Engand wird diese Art aus dem Hettangium und Unter-Sinemurium beschrieben, HARLOFF (1993) bezeichnet eine aus dem Unter-Pliensbachium der Schwäbischen Alb stammende ähnliche Art mit Cytherella sp. A, und nach BRAND (1990) kommt die Art noch im Ober-Bathonium von Nordwestdeutschland vor. In der Langenbrückener Senke, im Randen/Zürcher Weinland und im Wutachgebiet setzt diese Art jedoch erst im höheren Ober-Sinemurium ein, in Profil Aselfingen reicht sie bis ins Unter-Pliensbachium hinein (BEHER, 2004). Im Bereich der mittleren Schwäbischen Alb und in der Nordschweiz ist sie auf das tiefere Ober-Sinemurium beschränkt.
4. Fabalicypris praelongaDONZE, 1966: In Frankreich ist diese Art von Hettangium bis ins tiefe Sinemurium hinein verbreitet (DONZE, 1966, 1985). In Süddeutschland und Nordschweiz ist sie überwiegend auf das tiefere Ober-Sinemurium beschränkt, einzig im Bereich der östlichen Schwäbischen Alb gibt es Funde aus der Grenze Ober-Sinemurium/Unter-Pliensbachium (BEHER, 2004).
5. Pontocyprella fabaeformis (DREXLER, 1958): BEHER (2004) beschreibt diese Art aus dem Sinemurium und Unter-Pliensbachium von Süddeutschland und Nordschweiz. Dies entspricht der bei HARLOFF (1993) angegebenen Verbreitung. Desweiteren wird sie aus dem Hettangium (RIEGRAF, 1985) und dem Ober-Pliensbachium von Nordeuropa (CHRISTTENSEN, 1968, MICHELSEN, 1975; SIVHED, 1980) beschrieben. Im Tethysraum kommt sie ebenfalls von Hettangium bis Ober-Pliensbachium vor (HARLOFF & JÄGER, 1994; JÄGER, 1997), auf Sizilien im höheren Ober-Pliensbachium (BARBIERI, 1964).
6. Die hier mit Ostracode 1 bis Ostracode 4 bezeichneten Arten sind bisher unbekannt und bei der biostratigraphischen Gliederung des Bohrprofils nicht hilfreich.
Eine eindeutige Gliederung der Gesteinsabfolge mit Ostracoden scheint nicht möglich. Cytherella concentrica kommt bereits an der Basis des Profils vor und reicht bis ca. 1,5 ⅿ unterhalb des Tops. Donzocythere convergens dagegen tritt in diesem Profil recht spät auf und ist auf eine Probe beschränkt. Daher dürfte die Grenze Ober-Sinemurium/Unter-Pliensbachium darüber zu finden sein. Ihre genaue Lage kann jedoch nicht bestimmt werden. Da Cytherelloidea lacertosa häufig bereits im Ober-Sinemurium angetroffen werden kann, könnte das Unter-Pliensbachium auch oberhalb der untersuchten Profilstrecke liegen.
4. Beschreibung einiger Ostracoden aus dem Molassebecken
Bei den hier angeführten Ostracoden handelt es sich um Formen, welche bisher in der Literatur nicht beschrieben worden sind. Ihr Vorkommen scheint sich auf das Molassebecken zu beschränken, ihre biostratigraphische Zugehörigkeit ist nicht bekannt. In den Bohrungen Harresheim 1 und Markdorf 3 wurde das Gesteinsmaterial als Lias beta und gamma eingestuft, wobei eine lithologische Trennung beider Gesteinsfolgen mit dem Gamma-Log-Verfahren nicht möglich ist.
Alle Ostracoden befinden sich in der Belegsammung zur Dissertation Beher (2001) und werden im Staatlichen Museum für Naturkunde Stuttgart aufbewahrt.
Ostracode 1
siehe Schautafel, Fig. 1
BLNr. MA/9⁄184, LK; Ⅼ: 0,74217 mm; H: 0,4366 mm.
Material:
160 Gehäuse, 33 rechte und 32 linke Klappen.
Vorkommen:
Sinemurium und Unter-Pliensbachium, Arietenkalk bis Numismalismergel; Molassebecken, [Profile: Harresheim, Markdorf.]
Beschreibung:
Die nahezu rechteckigen Gehäuse erinnern in Lateralansicht an die Gattung Kinkelinella MARTIN. Der Vorderrand ist gleichmäßig breit, der Hinterrand schief gerundet. Letzterer ist leicht herabgezogen, so daß der hinterste Punkt des Gehäuses fast auf der Ventrallinie liegt. Der vorderste Gehäusepunkt liegt dagegen auf halber Gehäusehöhe. Der Ventralrand ist schwach konvex und sein mittlerer Abschnitt wird durch die etwas aufgeblähten und überhängenden Lateralseiten verdeckt. Der Dorsalrand ist ebenfalls nur leicht konvex und wird durch deutliche Kardinalwinkel in drei Abschnitte geteilt. Im Gegensatz zu den bisher vorgestellten Kinkelinella-Arten kann im Bereich des vorderen Schlosses keine auffällige Ausbuchtung beobachtet werden. Nach hinten zu fallen der mittlere und hintere Abschnitt leicht ab, Dorsal‑ und Ventralrand konvergieren nach hinten zu. Die Gehäuseenden sind mit einem glatten, breiten Randsaum versehen, so daß sie in Dorsalansicht kielartig vorspringen. Eine Skulptur ist erhaltungsbedingt nur schwach ausgeprägt. Neben glattschaligen, an Progonocythere stilla SYLVESTER-BRADLY erinnernde Formen treten auch solch mit einer feinen Längsberippung auf. Diese Längsripppen breiten sich im Zentrum fächerartig aus und laufen zum Hinterende wieder zusammen. Dazwischen sind häufig feine Querrippen ausgebildet, die die Längsrippen untereinander verbinden. Besonders charakteristisch ist eine kreisförmige “Schlaufe”. Sie befindet sich in der Gehäusemitte unterhalb des Dorsalrandes und wird durch eine scharf abbiegende Längsrippe und eine dazwischen liegende Querrippe gebildet. Das Schloß ist merodont, die Schloßzähne sind im Vergleich zu Kinkelinella Martin klein und unscheinbar; Muskeleindrücke und andere Internstrukturen sind erhaltungsbedingt nicht zu erkennen.
Bemerkungen:
Nach der Faunenzusammensetzung der Profile dürfte diese Art im Unter-Sinemurium verbreitet sein. Wichtiger Anhaltspunkt hierzu ist die Verbreitung von Donzocythere convergens (DONZE), welche bisher nur aus dem Unter-Sinemurium und tiefsten Ober-Sinemurium bekannt ist. Im Ober-Sinemurium des behandelten Gebietes sind bisher keine ähnlichen Formen aufgetreten
Ostracode 2
siehe Schautafel, Fig. 3
BLNr. MA/9⁄181, G; Ⅼ: 0,5828 mm; H: 0,2822 mm.
Material:
10 Gehäuse und 1 rechte Klappe.
Vorkommen:
Ober-Sinemurium und Unter-Pliensbachium, Obtusutone und Numismalismergel; Molassebecken, [Profile: Harresheim, Markdorf.]
Beschreibung:
Die Gehäuse sind niedrig und langgestreckt. Der Dorsalrand ist gerade und weist deutliche Kardinalwinkel auf, er fällt schräg zum Hinterende zu ab. Der Ventralrand ist in seiner Mitte stark konkav. Der Vorderrand ist breit, der Hinterrand enger und etwas asymmetrisch gerundet. Vorder‑ und Hinterrand sind mit einem breiten, deutlich abgeflachten und glatten Randsaum versehen. Die Lateralflächen sind mäßig gewölbt und überragen den Bereich des hinteren Schlosses leicht. Sie werden von einer grobmaschigen Skulptur bedeckt. Am vorderen Schloß ist ein Augenknoten entwickelt.
Bemerkungen:
Im Ober-Sinemurium des behandelten Gebietes sind bisher keine ähnlichen Formen aufgetreten. Da die nicht näher bestimmbare Art zusammen mit Donzocythere concergens (DONZE), einer Art aus dem Unter-Sinemurium, vorkommt, dürfte die Schichtenfolge aus der dieses Material stammt, dem Unter-Sinemurium angehören.
Ostracode 3
siehe Schautafel, Fig. 4
BLNr. HA/4⁄177, G; H: 0,5271 mm; H: 0,2660 mm;
BLNr. MA/9⁄180, G; Ⅼ: 0,5103 mm; H: 0,2732.
Material:
4 Gehäuse.
Vorkommen:
Ober-Sinemurium und Unter-Pliensbachium, Obtusustone und Numismalismergel; Molassebecken, [Profil: Markdorf.]
Beschreibung:
Die plumpen Gehäuse sind im Umriß nahezu rechteckig. Der Dorsalrand verläuft gerade, im Bereich des vorderen Schlosses ist er leicht ausgebuchtet. Der Ventralrand ist bei kleineren Exemplaren gerade, bei größeren in der Mitte konkav. Vorder‑ und Hinterrand sind gleichmäßig breit gerundet.
Beide werden von einem breiten Randsaum begleitet. Die Schalenoberfläche zeigt knopfartige Erhebungen und eine grobmaschige Retikulation. Eine wulstartig verdickte Rippe zieht an Hinter‑ und Ventralrand entlang nach vorn und und begletet ebenfalls den Vorderrand bis auf die halbe Gehäusehöhe. Im Bereich des Dorsalrandes befinden sich drei Wülste: Der Erste befindet sich anterodorsal, der Zweite in einer schwachen Depression unterhalb des Vorderschlosses und der Dritte unterhalb des hinteren Schlosses. Da nur Gehäuse vorliegen, können über Schloß und Muskeleindrücke keine Angaben gemacht werden.
Bemerkungen:
Aus dem Ober-Sinemurium des Arbeitsgebiets ist keine ähnliche Art bekannt. Wegen ihres Auftretens zusammen mit Doncocythere convergens (DONZE) ist davon auszugehen, daß es sich bei hier untersuchten Schichtfolge nicht um Obtustone handelt.
Ostracode 4
siehe Schautafel, Fig. 5
BLNr. MA/9⁄183, G; Ⅼ: 0,5706 mm; H: 0,2577 mm;
BLNr. HA/4⁄178, G, Ⅼ: 0,5871 mm; H: 0,2921 mm.
Material:
83 Gehäuse, 8 rechte und 5 linke Klappen.
Vorkommen:
Ober-Sinemurium und Unter-Pliensbachium, Obtusutone und Numismalismergel; Molassebecken, [Profile: Harresheim, Markdorf.]
Beschreibung:
Im Habitus entsprechen die Klappen und Gehäuse der Gattung Kinkelinella MARTIN. Der Dorsalrand ist konvex und weist im Bereich des vorderen Schlosses die typische Ausbuchtung auf. Der Ventralrand ist gerade bis schwach konkav und sein mittlerer Abschnitt wird von den nur leicht überhängenden Lateralflächen verdeckt. Der Vorderrand ist gleichmäßig breit und der Hinterrand eng gerundet. Beide Gehäuseenden werden von einem glatten, breiten Randsaum begleitet. Deshalb springen sie in Dorsalansicht kielartig vor. Die Skulptur besteht aus einigen Längsrippen. Ein Rippenpaar zieht den Ventralrand entlang nach hinten, ohne den Randsaum je zu erreichen. Drei Rippen verlaufen über die Lateralfläche, wobei sie aber weit hinter dem Vorderrand beginnen, und nacheinander enden. Eine einzige bleibt übrig und trifft auf den hinteren Randsaum. Dort kehrt sie um und bildet so eine kleine Schlaufe. Ein ähnlicher Rippenverlauf kommt bei Kinkelinella (E.) laqueata (KLINGLER & NEUWEILER) vor. Eine weitere Rippe beginnt am Vorderende unterhalb der halben Gehäusehöhe und zieht schräg aufwärts nach dorsal. In der Mitte biegt sie in einem deutlichen Knick zum Hinterende hin um und endet vor Erreichen des hinteren Randsaumes. Die letzte Rippe entspringt hinter dem Schloßohr und verläuft scharf am Dorsalrand entlang nach hinten. Sie erreicht den Randsaum.
Im hinteren Klappenbereich ist die Rippe gratartig erhöht, sie bildet so eine scharfe Kante und ist deshalb auch bei schlecht erhaltenen Exemplaren gut sichtbar. Das Schloß ist merodont, Muskeleindrücke sind nicht erkennbar. Es kann ein ausgeprägter (Geschlechts‑) Dimorphismus beobachtet werden, wie er ähnlich bei Kinkelinella (Ektyphocythere) laqueata (KLINGLER & NEUWEILER) und Kinkelinella (K.) sulcata (KLINGLER & NEUWEILER) auftritt.
Bemerkungen:
Bei Ostracode 4 könnte es sich um eine Form der Gattung Kinkelinella handeln. Trotz der Gemeinsamkeiten mit Kinkelinella (Ektyphocythere) laqueata dürfte es sich nicht um diese Art handeln. Bei Kinkelinella (E.) laqueata ziehen einige Längsrippen zum Dorsalrand und enden dort, eine Dorsalrippe fehlt. Auch ist die für Kinkelinella (E.) laqueata so charakteristische Schleife im Zentrum der Lateralfläche bei dem untersuchten Material nicht vorhanden.
Wegen ihres Auftretens zusammen mit Doncocythere convergens (DONZE) dürfte die Art stratigraphisch in das Unter-Sinemurium gehören.
Alle Ostracoden befinden sich in der Belegsammung zur Dissertation BEHER (2001) und werden im Staatlichen Museum für Naturkunde Stuttgart aufbewahrt.
5. Literatur
AINSWORTH, N. R. (1989): Rhaetian, Hettangian and Sinemurian Ostracoda from the Fasnet Basin, offshore Southwest Ireland.- Geol. Surv. Ire. Bull., 4⁄2: 107–150, 13 5 Abb., 9 Tab., 4 Taf.; Dublin.
APOSTOLESCU, Ⅴ. (1959): Ostracodes du Lias du bassin de Paris.- Rév. Inst. Franc. Petrole, 14⁄6: 795–826, 4 Abb., 4 Taf., 1 Tab.; Paris.
BARBIERI, F. (1964): Micropaleontologia del Liea epsilon Dogger del poso Ragusa 1 (Sicilia).- Riv. Ital. Paleont., 19⁄4: 709–830, Taf. 56–64; Milano.
BEHER , E. (2001): Biostratigraphie mit Ostracoden im Ober-Sinemurium von Süddeutschland und Nordschweiz.- Diss. Univ. Heidelberg, 278 S., 54 Abb., 17 Taf., 1 Tab.; Heidelberg. Manuskript.
BEHER, E. (2004): Ostracodenfauna und Biostratigraphie im Ober-Sinemurium von Süddeutschland und der Nordschweiz.- Stuttgarter Beitr. Naturk., Ser. B., 349: 171 S., 28 Abb., 16 Taf.; Stuttgart.
BRAND, E. (1990): Biostratigraphische Untergliederung des Ober-Bathonium im Raum Hildesheim, Dordwestdeutschland mittels Ostracoden und Korrelation ihrer Vertikalreichweite mit Ammonitenzonen.- Geol. Jb., A 121: 119–273, 4 Abb., 1 Tab., 18 Taf.; Hannover.
CHRISTENSEN; O. B. (1968): Biostratigraphical investigation of the Pre-Quaternary in the Øresund Boreholes-mainly on the basis of ostracods.- In: LARSEN, G., CHRISTENSEN, O. B., BAY, I. & BUCH, A.(1968): Øresund. HelsngØr-Hälsingborg Linien, Geologisk Rapport.- Danmark geol. Unders., Rapp. 1: 81–85, Tab. 23; KØbenhavn.
CONTINI, D. & PARIWATVORN, P. (1964): Etude comparée de la microfaune du Lias supérieur d’Aselfingen (Jura du Randen) et de Pouilley-les-Vignes (Jura Franc-Contois).- Annales scientif. de l’Univ. de Besançon, 2 sér., Géologie, fasc. 18: 35–44, 8 Abb.; Besançon.
DONZE, P. (1966): Ostracodes de l’Hettangien entre Aubenas et Privas (Ardèche).- Trav. Lab. géol. Fac. Sci. Lyon, n. sèr. 13: 121–139, Taf. 5–7; Lyon.
DONZE, P. (1968): Especes nouvelles d’ostracodes du Lias inférieur Normand.- Bull. Soc. Linn. Normandie, 9: 78–87, 2 Taf.; Caen.
DONZE, P. (1985): Lias inférieur et moyen.- In: OERTLI, H. J. (Hrsg.): Atlas des Ostracodes de France, Mém. Elf Aquitane, 9: 101–117, Taf. 21–26, Abb. 9–14, Tab. 5; Pau.
HARLOFF, J. (1993): Ostracoden des Unter-Pliensbachiums in Baden‑ Württember.- Stuttgarter Beitr. Naturk., Ser. B, 191: 1–214, 75 Abb., 9 Tab., 14 Taf.; Stuttgart.
HARLOFF, J. & JÄGER, R. (1994): Ostrakoden aus dem Lias der Kalkalpen Bayerns und Nordtirol.- Stuttgarter Beitr. Naturk., Ser. B., 205: 63 S., 4 Abb., 8 Taf.; Stuttgart.
JÄGER, R. J. (1997): Foraminiferen und Ostracoden aus den Fleckenmergeln des kalkalpinen Lias.- Münchner Geowiss. Abh., Reihe A, 33: ,1–111, 36 Abb., 15 Tab., 5 Taf.; München.
MICHELSEN, O. (1975): Lower Jurassic stratigraphy and Ostracoda of the Danish Embayment.- Dan. Geol. Unders. Ⅱ Raekke, 94: 1–60, 45 Abb., 13 Tab., 42 Taf.;Copenhaven. OERTLI, H. J. (1963): Faunes d’Ostracodes du mésozoique de France.- VIII+57 S., 90 Taf., 3 Abb., 4 Tab., 2 Kt; Leiden (Brill).
RIEGRAF, W. (1985): Mikrofauna, Biostratigraphie und Fazies im Unteren Toarcium Südwestdeutschlands und Vergleiche mit benachbarten Gebieten .- Tübinger mikropaläont. Mitt., 3: 232 S, 12 Taf., 33 Abb.; Tübingen.
SIVHED, U. (1980): Lower Jurassic Ostracodes and Statigraphy of Western Skåne, Southern Sweden.- Avh. Sver. Geol. Unders., Ca 50: 1–85, 40 Abb., 12 Taf.; Uppsala.
SCHNEIDER, J. (1962): Der Jura in Erdölbohrungen des westlichen Molassetroges.- Aldinger Festschrift (HRSG.: GEYER, O. F. & GWINNER, Ⅿ. P.), S 163–172, 1. Abb., 3 Beil.; Stuttgart.
WALTSCHEW, A. (2000): Die Mikrofauna des fränkischen Carixium (Unteres Pliensbachium, Lias).- Erlanger geol. Abh., 132: 91 S., 13 Abb., 4 Taf., 5 Tab.; Erlangen.